1.はじめに
現在、高コレステロール血症、動脈硬化症予防・改善のための多くの医薬品・特定保健用食品の登場、社会的関心とは裏腹に、世界保健機関(WHO)の統計によると、世界の死因の第1位は依然として心臓血管疾患であり、現在でも決定的な動脈硬化症の解決策は残念ながらない、というのも厳然とした事実である。高コレステロール血症制圧は動脈硬化症制圧につながることから、食物繊維、大豆たんぱく質などが研究されてきた。しかし満足できる成分が発見されていないことは、上記の事実からも明白ある。つまり、従来から世界中で研究されてきた大豆たんぱく質や食物繊維などの従来の食品や医薬品では、体内に余分に蓄積したコレステロールや摂取したコレステロールを超効率的に体外排出させることや、高コレステロール血症の治療は充分でない。また、そのための理論・技術も未成熟である。従って、コレステロール代謝を改善するための革新的な理論・技術が切望されている。
2.血清コレステロールと食品たんぱく質・ペプチド
高脂血症、特に高コレステロール血症は心臓病、特に、虚血性心疾患の最も重要な危険因子の一つと考えられている。事実わが国では、冠状動脈硬化症に基づく狭心症や心筋梗塞などの虚血性心疾患による死亡率の増加が著しい。このような背景から、食餌による高コレステロール血症の予防や改善も非常に有効な方策と考えられている。すなわち、食餌たんぱく質も血清コレステロール濃度調節因子として、かなり有用な成分である。図1のように、一般的に動物性たんぱく質に比べて、植物性たんぱく質の摂取によって血清コレステロールは低値を示すことが、これまで繰り返し語られてきた(参考論文1)。しかし、すでにわれわれは、動物性食品たんぱく質である牛乳乳清たんぱく質の血清コレステロール低下作用に関して研究してきている。詳細は既報(2〜5)を参照いただきたいが、驚くべきことに、牛乳乳清たんぱく質は従来、血清コレステロール低下作用を有することが明らかにされてきた大豆たんぱく質(6、7)よりも血清コレステロール低下の作用を大幅に促すことが確認された。
| 図1 |
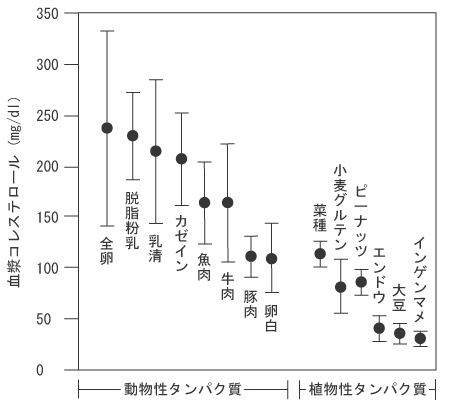 |
血清コレステロールを低下させるたんぱく質に関する研究は、90年以上前から行われてきたが、90年以上の長い間誰も発見できなかったコレステロール代謝改善ペプチド(IIAEK:牛乳から発見したことから「ラクトスタチン」(lactostatin)と命名)を、われわれは世界で初めて、乳清たんぱく質から発見した。(8)しかも、驚くべきことに、その作用は医薬品のβ-シトステロール(植物ステロール)よりも強力であった。(畜産の情報(国内編)2002年12月号参照)
これまでのところ、オリゴペプチドの媒介するコレステロール代謝調節系に関する報告はない。ラクトスタチンの標的とする遺伝子を特定することは、その遺伝子に作用する新しいコレステロール代謝改善作用を発揮するペプチドの発見に発展する可能性があり、たいへん興味深い。
そこで、この研究では、マウスを用いて、ラクトスタチンの摂取より、さまざまなコレステロール代謝関連遺伝子発現に対する影響を詳細に解析し、ラクトスタチンなどのオリゴペプチドの媒介する新しいコレステロール代謝調節系の解明をするため、下記の2点について検討することを目的とした。
(1)Caco-2培養細胞で発見した、コレステロール吸収抑制ペプチドが、in vivo(生体内)で血清コレステロール低下作用を有するかどうかをマウスに食餌として与えて評価検討した。これまでは、ラットに経口投与で与えた実験系で評価したため、ラットとは別の種においても活性が観察されるかどうかを検討した。そのために、短期摂取による摂取条件をマウスで設定した。
今回マウスを用いた動物実験系を構築する理由は、マウスは遺伝的な背景の解析がラットよりも進んでいる点、高価なペプチドの摂取・評価を効率的に展開できる点が有用であるためである。
(2)遺伝的な背景が既知であるマウスにおいて、ラクトスタチンのコレステロール代謝改善作用の発現機構を解明する目的で、コレステロール代謝の主要な臓器である肝臓のコレステロール代謝関連遺伝子などに与える影響を評価した。また、この標的遺伝子が特定されれば、この標的遺伝子の発現に影響するペプチドを探索評価することにより、より活性の高いオリゴペプチドが発見できる。
つまり、この標的遺伝子の特定結果は、ラクトスタチン以外の革新的な活性オリゴペプチドによる新しい効率的スクリーニングに道を拓くものであり、大いに期待できる。
3.研究内容
[実験]
動物実験(マウス)による新しいコレステロール代謝改善ペプチド(ラクトスタチン)のコレステロール代謝に対する影響およびラクトスタチンの肝臓コレステロール代謝関連遺伝子発現に対する影響の検討
実験動物として、初体重18〜20gのICR系雄マウス(4週齢)を用いた。コントロール群のマウスにはカゼイン20%とコレステロール1%を含む食餌を与え、ラクトスタチン群のマウスにはカゼイン20%とコレステロール1%とラクトスタチン0.14%を含む食餌を与え、それぞれ4日間飼育した。解剖前に4時間絶食して、心臓採血によりマウスをと殺し、血清コレステロールを測定した。また、肝臓脂質の測定も行った。さらに、肝臓から全RNAを抽出し、リアルタイム定量PCR法(ABIのTaqMan
probeを活用)により、コレステロール代謝関連遺伝子のmRNA定量を行った。それぞれの全RNAは群ごとに一つにまとめて測定した。
[統計分析]
実験結果の統計的分析には、Student's t-Test[注1]を用いた。
4.結果及び考察
(1)ラクトスタチンの血清コレステロール低下作用
ラクトスタチンが強力な血清コレステロール低下作用を有することは、以前ラットを用いて報告した(8)。しかし、マウスを用いた本研究においてはラクトスタチンの食餌への添加量が少なかったためか、有意な血清コレステロール低下作用は見られなかったが、血清コレステロールの低下傾向は観察できた(表1)。
| 表1 C(対照)、ラクトスタチンを与えたマウスにおける体重増加量、食餌摂取量、肝臓重量、血清コレステロール、動脈硬化指数 |
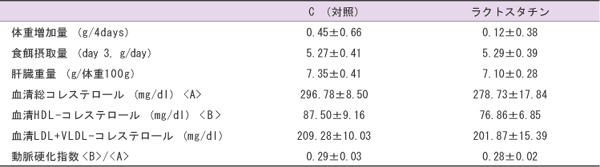 |
| 注:値は各群8匹の平均値±標準誤差で示した。 |
(2)ラクトスタチンの肝臓脂質、コレステロールプール低下作用
本研究において、ラクトスタチンを与えた群では、全ての肝臓脂質及びコレステロールプールで低下傾向が見られた(表2)。コレステロールプールは血清総コレステロールと肝臓総コレステロールの和で示されるが、大半は肝臓総コレステロールが占める。そのため、ラクトスタチンの肝臓総コレステロール低下傾向は非常に大きな意味をもつ。これは血清コレステロールと同様、有意な差は見られなかったものの、ラクトスタチンの添加量をさらに増加させることで有意な効果が期待できる可能性がある。
| 表2 C (対照)、ラクトスタチンを与えたマウスにおける肝臓脂質とコレステロールプールC (対照)ラクトスタチン |
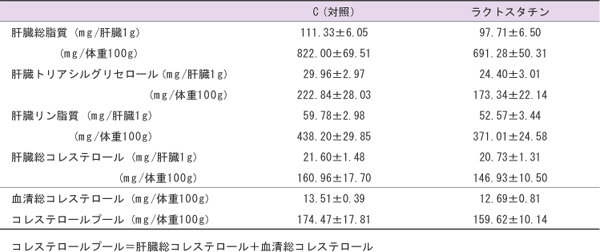 |
| 注:値は各群8匹の平均値±標準誤差で示した。 |
(3)ラクトスタチンの肝臓コレステロール代謝関連遺伝子のmRNA量に対する影響
各遺伝子についての値はβ-アクチンの値で補正し、コントロール群の値を100としてラクトスタチン群の値を相対的に示した(図2)。まずラクトスタチン群では、胆汁酸合成の律速酵素[注2]であるコレステロール7α-水酸化酵素(CYP7A1)が約66%も減少した。CYP7A1の発現に関係する遺伝子のうち、mRNAを定量したものは転写因子であるSHP-1、FTF、FXR、LXRαである。CYP7A1の発現抑制経路として、FXRが胆汁酸により活性化されることでFTFが増加し、さらにそのFTFがSHP-1およびCYP7A1を増加させるが、SHP-1はFTFと結合してSHP-1自身およびCYP7A1を減少させるという経路が報告されており(9)、ほかにも同様の報告がある(10〜13)。また、CYP7A1の発現促進経路として、コレステロールから生成したオキシステロールにより活性化されたLXRαがCYP7A1を増加させ、胆汁酸合成系を促進するという経路が報告されている(12〜14)。しかし、ラクトスタチンによりCYP7A1は減少したにもかかわらず、LXRαは変化が無かった。また、FXRは減少しているのにもかかわらず、FTFが増加した。そして、SHP-1は大きく減少したにもかかわらず、CYP7A1が減少した。以上のように、本研究のmRNA定量結果は過去の報告に反する結果となった。また、同じく胆汁酸合成に関与する酵素であるCYP8B1については、CYP7A1と同様の調節機構が報告されているが(10、11、15、16)、CYP7A1とは全く異なった挙動を示した。従って、ラクトスタチンのCYP7A1やCYP8B1への影響については、今後詳細な検討が必要である。またコレステロール合成系の律速酵素であるHMG-CoA
還元酵素が減少した。コレステロールプールが減少傾向を示したのはこのためである可能性が考えられる。
| 図2 C(対照)、ラクトスタチンを与えたマウスにおける肝臓コレステロール代謝関連遺伝子のmRNA量(β-actinで補正) |
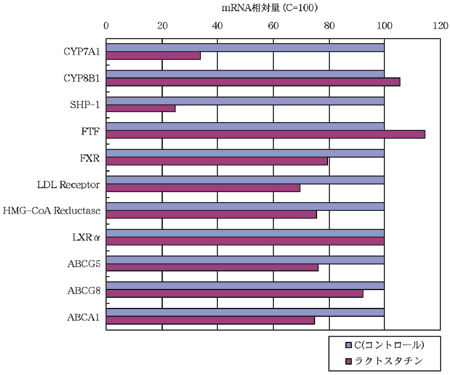 |
次に、LDL(低密度リボタンパク質)受容体について、CYP7A1が減少したことにより胆汁酸合成量が減少し、コレステロール吸収量が減少したことで、肝臓におけるLDL受容体の必要量が低下した可能性も考えられる。またコレステロールの輸送や搬出(排せつ・吸収)と関連することが知られているABCトランスポーターであるABCG5、ABCG8、ABCA1は胆汁へのコレステロールの排せつを促すという報告があるが(17、18)、本研究ではコレステロールプールが減少傾向を示しているのにも関わらず、ABCG5、ABCG8、ABCA1は全て減少した。さらに、ABCG5、ABCG8、ABCA1を活性化するLXRα(17、19)に変化が無いことから、ラクトスタチンのコレステロールプール低下傾向はこれらの肝臓ABCトランスポーターとの関連性が薄い可能性が考えられる。
従って、ラクトスタチンは胆汁酸生合成の律速酵素であるコレステロール7α-水酸化酵素のmRNAを抑制し、コレステロール吸収を抑制するとともに、コレステロールの吸収排せつを促進することにより、コレステロール代謝改善作用を発揮する可能性が示唆された。これらの成果は、世界初のコレステロール代謝改善ペプチド(ラクトスタチン)の標的遺伝子を世界で初めて特定した成果である。この標的遺伝子の特定結果は、ラクトスタチン以外の革新的な活性オリゴペプチドの新しい効率的スクリーニングに道を拓くものであり、大いに期待できる。今後さらに詳細に検討していく必要がある。
ところで、現代日本の欧米型の食生活への移行は、身体の向上や栄養の改善をもたらしたが、その一方で、肥満、高脂質血症、高コレステロール血症などの増加に伴う、動脈硬化症などの心臓血管疾患すなわち、生活習慣病の増加が指摘されている。これらの生活習慣病増加の原因をともすれば、動物性食品すなわち、卵・肉・牛乳などの畜産物に求める傾向があることも事実である。しかし、本研究で明らかにしたように、牛乳成分には、高コレステロール血症などの生活習慣病を予防改善するラクトスタチンなどの有用ペプチドが存在するわけであり、今後の牛乳の付加価値製品の開発に少しでも寄与することを期待したい。
参考論文
1.Carrol, K. K. and Hamilton, R. M. G. (1975) Effects of dietary protein
and carbohydrate on plasma cholesterol levels in relation to atherosclerosis.
J.Food Sci., 40, 18-23.
2.Nagaoka, S., Kanamaru, Y. and Kuzuya,Y. (1991) Effects of whey protein
and casein on the plasma and liver lipids in rats. Agric.Biol.Chem., 55,
813-818.
3.Nagaoka, S., Kanamaru, Y., Kuzuya, Y., Kojima, T and Kuwata, T. (1992)
Comparative studies on the serum cholesterol lowering action of whey protein
and soybean protein in rats.
Biosci.Biotech.Biochem., 56, 1484-1485.
4.長岡 利、金丸義敬、葛谷泰雄:乳清タンパク質の血清コレステロール低下作用、山内邦男、今村経明、守田哲朗 責任編集"牛乳成分の特性と健康"東京:光生館、57-84
(1993)
5.長岡 利:食品成分および生体異物によるコレステロール代謝制御に関する研究(日本栄養食糧学会奨励賞受賞総説)日本栄養食糧学会誌、49, 303-313(1996)
6.Nagata, Y., Ishiwaki, N and Sugano, M. (1982) Studies on the mechanisms
of antihypercholesterolemic action of soy protein and soy protein-type
amino mixtures in relation to the casein counterparts in rats. J.Nutr.,
112, 1614-1625.
7.Nagaoka, S., Awano, T., Nagata, N., Masaoka, M., Hori, G. and Hashimoto,
K. (1997) Serum cholesterol reduction and cholesterol absorption inhibition
in CaCo-2 cells by a soyprotein peptic hydrolyzate. Biosci.Biotech.Biochem.,
61, 354-356.
8.Nagaoka, S., Futamura, Y., Miwa, K., Awano, T., Yamauchi, K., Kanamaru,
Y., Kojima, T., and Kuwata, T. (2001) Identification of novel hypocholesterolemic
peptides derived from bovine milk β-lactoglobulin. Biochem Biophys Res
Commun 281, 11-17.
9.Chen, W., Owsley E., Yang, Y., Stroup, D. and Chiang, J. Y. (2001) Nuclear
receptor-mediated repression of human cholesterol 7alpha-hydroxylase gene
transcription by bile acids. J. Lipid Res., 42, 1402 -1412.
10.Lee, Y. K. and Moore, D. D. (2002) Dual mechanisms for repression of
the monomeric orphan receptor liver receptor homologous protein-1 by the
orphan small heterodimer partner. J. Biol. Chem., 277, 2463-2467.
11.Goodwin, B., Jones, S.A., Price, R.R., Watson, M.A., McKee, D.D., Moore,
L.B., Galardi, C., Wilson, J.G., Lewis, M.C., Roth, M.E., Maloney, P.R.,
Willson, T.M. and Kliewer, S.A. (2000) A regulatory cascade of the nuclear
receptors FXR, SHP-1, and LRH-1 represses bile acid biosynthesis. Mol.
Cell, 6, 517-526.
12.Lu, T.T., Makishima, M., Repa, J.J., Schoonjans, K., Kerr, T.A., Auwerx,
J. and Mangelsdorf, D.J. (2000) Molecular basis for feedback regulation
of bile acid synthesis by nuclear receptors. Mol. Cell, 6, 507-515.
13.Davis, R.A., Miyake, J.H., Hui, T.Y. and Spann, N.J. (2002) Regulation
of cholesterol-7α-hydroxylase: BAREly missing a SHP. J. Lipid Res., 43,
533-543.
14.Peet, D. J., Turley, S. D., Ma, W., Janowski, B. A., Lobaccaro, J.
M. A., Hammer, R. E. and Mangelsdorf, D. J. (1998) Cholesterol and bile
acid metabolism are impaired in mice lacking the nuclear oxysterol receptor
LXR alpha. Cell, 93, 693-704.
15.Castillo-Olivares, A. and Gil, G. (2000) Alpha 1-fetoprotein transcription
factor is required for the expression of sterol 12alpha-hydroxylase, the
specific enzyme for cholic acid synthesis. Potential role in the bile
acid-mediated regulation of gene transcription. J. Biol. Chem., 275, 17793-17799.
16.Castillo-Olivares, A. and Gil, G. (2001) Suppression of sterol 12alpha-hydroxylase
transcription by the short heterodimer partner: insights into the repression
mechanism. Nucleic acids Res., 29, 4035-4042.
17.Plosch, T., Kok, T., Bloks, V. W., Smit, M. J., Havinga, R., Chimini,
G., Groen, A. K. and Kuipers, F. (2002) Increased hepatobiliary and fecal
cholesterol excretion upon activation of the liver X receptor is independent
of ABCA1. J. Biol. Chem., 277, 33870 -33877.
18.Yu, L., Li-Hawkins, J., Hammer, R. E., Berge, K. E., Horton, J. D.,
Cohen, J. C. and Hobbs, H. H. (2002) Overexpression of ABCG5 and ABCG8
promotes biliary cholesterol secretion and reduces fractional absorption
of dietary cholesterol. J. Clin. Invest., 110, 671-680.
19.Repa, J. J., Berge, K. E., Pomajzl, C., Richardson, J. A., Hobbs, H.
and Mangelsdorf, D. J. (2002) Regulation of ATP-binding cassette sterol
transporters ABCG5 and ABCG8 by the liver X receptors alpha and beta.
J. Biol. Chem., 277, 18793 -18800.
[注1]実験データの統計処理方法の1つで、2群の実験群間での統計的な有為差を検討するために頻用される。
[注2]代謝径路において、一連の酵素反応により反応が進行するときにその代謝の速度や方向を支配調節する酵素のことをいう。