仢挷嵏丒曬崘
抺嶻暔廀梫奐敪挷嵏尋媶偐傜
僐儗僗僥儘乕儖戙幱傪夵慞偡傞崅晅壛壙抣媿擕奐敪偺偨傔偺婎慴尋媶
亅媿擕偺怴偟偄寣惔僐儗僗僥儘乕儖掅尭壔儁僾僠僪偵娭偡傞尋媶亅
婒晫戝妛丂擾妛晹丂
彆嫵庼丂挿壀丂棙
偼偠傔偵
丂尰戙擔杮偺墷暷宆偺怘惗妶傊偺堏峴偼丄恎懱偺岦忋傗塰梴偺夵慞傪傕偨傜偟
偨偑丄偦偺堦曽偱丄旍枮丄崅帀幙寣徢丄崅僐儗僗僥儘乕儖寣徢側偳偺憹壛偵敽
偆丄摦柆峝壔徢側偳偺怱憻寣娗幘姵偡側傢偪丄惗妶廗姷昦偺憹壛偑巜揈偝傟偰
偄傞丅偙傟傜偺惗妶廗姷昦偺憹壛偺尨場傪偲傕偡傟偽丄摦暔惈怘昳偡側傢偪丄
棏丒擏丒媿擕側偳偺抺嶻暔偵媮傔傞孹岦偑偁傞偙偲傕帠幚偱偁傞丅偮傑傝丄摦
暔惈怘昳偼惗妶廗姷昦偲偺娭楢偐傜宧墦偝傟偑偪偱偁傞丅杮摉偵丄摦暔惈怘昳
偺愛庢偼寬峃偵岲傑偟偔側偄偙偲偽偐傝側偺偩傠偆偐丅妋偐偵丄恾1偺傛偆偵丄
摦暔惈帀朾偺愛庢検偺懡偄崙柉偼丄怱嬝峓嵡偺巰朣棪偑崅偄偙偲偑曬崘偝傟偰
偄傞丅
丂偟偐偟丄媿擕側偳偺摦暔惈僞儞僷僋幙偼怉暔惈僞儞僷僋幙偲斾妑偟偰丄堦斒
偵僶儔儞僗偺偲傟偨傾儈僲巁慻惉傪桳偟偰偍傝丄崅偄傾儈僲巁僗僐傾傪帵偡偙
偲傕抦傜傟偰偄傞丅幚嵺偵恾2偺傛偆偵丄擔杮偵偍偄偰偼丄棏丄媿擕丄擏偺徚
旓検偺憹壛偲暯嬒庻柦偺怢傃偑枾愙偵娭楢偟偰偄傞偙偲傕曬崘偝傟偰偄傞丅偙
偺梫場偼丄堛椕傗惗妶娐嫬偺岦忋傕栜榑偱偁傠偆偑丄怘惗妶丄摿偵摦暔惈怘昳
偺幙丒検偺岦忋偑戝偒偔婑梌偟偰偄傞偲峫偊傜傟傛偆丅
丂埲忋偺傛偆側憡柕弬偟偨曬崘偑側偝傟偰偄傞偙偲偐傜丄巹偼摦暔惈怘昳偺桳
岠惈偵娭偡傞恀幚偺巔偵敆傝偨偄偲偄偆偙偲傪偐偹偑偹峫偊偰偍傝丄摦暔惈怘
昳惉暘偺昡壙偵嫮偄娭怱偲嫽枴傪書偄偰偒偨傢偗偱偁傞丅
仦恾1丗擏椶丒寋棏丒擕惢昳偺徚旓検偲暯嬒庻柦偺曄壔仦
仦恾俀丗悽奅奺崙偺屌宍乮摦暔惈乯帀朾偲塼忬乮怉暔惈乯帀朾偺愛庢忬嫷偲怱嬝峓嵡巰朣棪偲偺娭學仦
愜傟慄僌儔僼丗怱嬝峓嵡巰朣幰妱崌乮抝巕乯
朹僌儔僼丗塼忬帀朾乮敀乯偲屌宍帀朾乮椢怓乯
乮徏嶈弐媣亀榁壔偲塰梴亁乮幝尨丒怷撪丒嵶扟曇乯戞堦弌斉乮1988乯傛傝夵曄乯
寣惔僐儗僗僥儘乕儖偲怘昳僞儞僷僋幙丒儁僾僠僪
丂崅帀寣徢丄摿偵崅僐儗僗僥儘乕儖寣徢偼怱憻昦丄摿偵嫊寣惈怱幘姵偺嵟傕廳
梫側婋尟場巕偺1偮偲峫偊傜傟偰偄傞丅帠幚傢偑崙偱偼丄姤忬摦柆峝壔徢偵婎
偯偔嫹怱徢傗怱嬝峓嵡側偳偺嫊寣惈怱幘姵偵傛傞巰朣棪偺憹壛偑挊偟偄丅偙偺
傛偆側攚宨偐傜丄崅僐儗僗僥儘乕儖寣徢偺怘塧偵傛傞梊杊傗夵慞傕旕忢偵桳岠
側曽嶔偲峫偊傜傟偰偄傞丅偡側傢偪丄怘塧僞儞僷僋幙傕寣惔僐儗僗僥儘乕儖擹
搙挷愡場巕偲偟偰丄偐側傝桳梡側惉暘偱偁傞丅恾俁偺傛偆偵丄堦斒揑偵摦暔惈
偨傫傁偔幙偵斾傋偰丄怉暔惈偨傫傁偔幙偱寣惔僐儗僗僥儘乕儖偼掅抣傪帵偡偙
偲偑丄偙傟傑偱孞傝曉偟岅傜傟偰偒偨乮嶲峫榑暥1乯丅偟偐偟丄婛偵傢傟傢傟
偼丄摦暔惈怘昳偨傫傁偔幙偱偁傞媿擕擕惔偨傫傁偔幙偺寣惔僐儗僗僥儘乕儖掅
壓嶌梡偵娭偟偰尋媶偟偰偒偰偄傞丅徻嵶偼婛曬乮2乣5乯傪嶲徠偄偨偩偒偨偄偑丄
嬃偔傋偒帠偵丄廬棃寣惔僐儗僗僥儘乕儖掅壓嶌梡傪桳偡傞偙偲偑柧傜偐偵偝傟
偰偒偨戝摛偨傫傁偔幙乮6丄7乯傛傝傕嫮椡側寣惔僐儗僗僥儘乕儖掅壓嶌梡傪媿
擕擕惔偨傫傁偔幙偑敪尰偡傞偙偲傪敪尒偟偨丅偝傜偵丄傢傟傢傟偺尋媶幒偱偼丄
偙傟傑偱偵媿擕偺擕惔僞儞僷僋幙偱偁傞兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛
悈暘夝暔乮LTH乯偑丄僐儗僗僥儘乕儖戙幱夵慞嶌梡傪桳偡傞偙偲傕曬崘偟偰偒
偨乮5乯丅偟偐傕丄偙偺嶌梡偼戝摛僞儞僷僋幙傛傝傕偼傞偐偵嫮椡偱婜懸偱偒
傞丅偝傜偵LTH傪暘夋暘庢偟偰丄彫挵忋旂嵶朎偺婡擻傪敪尰偡傞Caco-2嵶朎偵
偍偗傞僐儗僗僥儘乕儖偺媧廂昡壙宯偵傛傝丄媧廂梷惂嶌梡傪桳偡傞儁僾僠僪偺
摨掕傪峴偭偨丅偦偺寢壥丄IIAEK丄GLDIQK丄ALPMH丄VYVEELKPTPEGDLEILLQK偺4
庬椶偺怴偟偄僐儗僗僥儘乕儖媧廂梷惂儁僾僠僪傪敪尒偟偨乮8乯丅偦偺偆偪丄
偡偱偵IIAEK偼儔僢僩偵偍偄偰寣惔僐儗僗僥儘乕儖傪堛栻昳偺兝-僔僩僗僥儘乕
儖傛傝傕嫮椡偵掅壓偝偣傞偙偲傕悽奅偵愭嬱偗偰敪尒偟曬崘偟偨乮8乯丅
仦恾俁仦
丂偦偙偱丄杮憤愢偱偼丄兝-儔僋僩僌儘僽儕儞桼棃儁僾僠僪偺僐儗僗僥儘乕儖
戙幱夵慞嶌梡偵偮偄偰丄壓婰偺傛偆側3揰偵偮偄偰専摙偟偨惉壥傪徯夘偡傞丅
拲乯忋婰偺儁僾僠僪偺偆偪丄偦傟偧傟丄A丗傾儔僯儞丄D丗傾僗僷儔僊儞巁丄E
丗僌儖僞儈儞巁丄G丗僌儕僔儞丄H丗僸僗僠僕儞丄I丗僀僜儘僀僔儞丄K丗儕僔儞丄
L丗儘僀僔儞丄M丗儊僠僆僯儞丄P丗僾儘儕儞丄Q丗僌儖僞儈儞丄T丗僩儗僆僯儞丄
V丗僶儕儞丄Y丗僠儘僔儞偺傾儈僲巁傪帵偡丅
嘆丂Caco-2攟梴嵶朎偱敪尒偟偨丄僐儗僗僥儘乕儖媧廂梷惂儁僾僠僪偑丄in vi
丂vo乮懱撪偱乯偱寣惔僐儗僗僥儘乕儖掅壓嶌梡傪桳偡傞偐偳偆偐傪儅僂僗偵怘
丂塧偲偟偰梌偊偰昡壙専摙偟偨丅偙傟傑偱偼丄儔僢僩偵宱岥搳梌偱梌偊偨幚尡
丂宯偱昡壙偟偨偨傔丄儔僢僩偲偼暿偺庬偵偍偄偰傕妶惈偑娤嶡偝傟傞偐偳偆偐
丂傪専摙偟偨丅偦偺偨傔偵丄抁婜愛庢偵傛傞愛庢忦審傪儅僂僗偱愝掕偟偨丅崱
丂夞儅僂僗傪梡偄偨摦暔幚尡宯傪峔抸偡傞棟桼偼丄儅僂僗偼堚揱揑側攚宨偺夝
丂愅偑儔僢僩傛傝傕恑傫偱偄傞揰丄崅壙側儁僾僠僪偺愛庢丒昡壙傪岠棪揑偵揥
丂奐偱偒傞揰偑桳梡偱偁傞偨傔偱偁傞丅
嘇丂儅僂僗偺応崌傕丄儔僢僩偺応崌偲摨條偵丄IIAEK儁僾僠僪偼堛栻昳偱偁傞兝
丂-僔僩僗僥儘乕儖偲斾妑偟偰丄僐儗僗僥儘乕儖戙幱夵慞嶌梡傪傛傝嫮椡偵敪
丂尰偡傞偐偳偆偐傪斾妑専摙偟偨丅
嘊丂IIAEK埲奜偺丄in vitro乮懱奜偱乯偱僐儗僗僥儘乕儖媧廂傪梷惂偟偨GLDIQK丄
丂ALPMH儁僾僠僪偺僐儗僗僥儘乕儖戙幱偵懳偡傞塭嬁偵偮偄偰丄儅僂僗偱専摙偟
丂偨丅
尋媶撪梕
幚尡1
丂摦暔幚尡乮儅僂僗乯偵傛傞兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼
棃偺儁僾僠僪乮IIAEK乯偍傛傃堛栻昳兝-僔僩僗僥儘乕儖偺僐儗僗僥儘乕儖戙
幱偵懳偡傞塭嬁
丂ICR宯梇儅僂僗乮弶懱廳栺20g乯傪梡偄偨丅20亾僇僛僀儞偵1亾偺僐儗僗僥儘
乕儖傪揧壛偟偨帞椏偵丄僇僛僀儞僩儕僾僔儞壛悈暘夝暔乮CTH乯丄IIAEK偍傛
傃兝-僔僩僗僥儘乕儖傪偦傟偧傟0.3亾揧壛偟偨怘塧傪4擔娫愛庢偝偣偨丅怘塧
慻惉偼昞1偵帵偟偨丅4擔栚偵儔僢僩傪怱憻嵦寣偵傛傝偲嶦偟偨丅奺孮偼偦傟
偧傟6旵偲偟偨丅懱廳偼枅擔丄24帪娫偺怘塧愛庢検偼丄4擔栚偵應掕偟丄傆傫
偺嵦庢偼2擔栚偐傜4擔栚偵峴偭偨丅4擔栚偵儔僢僩傪僄乕僥儖杻悓壓偱怱憻嵦
寣偵傛傝偲嶦偟丄娞憻傪揈弌偟偨丅寣惔偼墦怱暘棧乮3000夞揮丄15暘乯偡傞
偙偲偵傛傝挷惢偟丄暘愅傑偱搥寢曐懚偟偨丅寣惔帀幙偼峺慺朄偱應掕偟偨丅
娞憻偼昞柺偵晅偄偨寣塼側偳傪傛偔傆偒庢傝丄0.9亾NaCl梟塼偱愻忩屻丄幖廳
検傪應掕偟丄暘愅傑偱搥寢曐懚偟偨丅娞憻帀幙偺拪弌丄惛惢偼丄Folch傜偺曽
朄乮9乯偵廬偭偰峴偄峺慺朄偱應掕偟偨丅傆傫拞僗僥儘僀僪攔煏検偼婛曬乮10乯
偵廬偭偰掕検偟偨丅
幚尡孮偼壓婰偺傛偆側傕偺偱偁傞丅
嘆CTH丗僇僛僀儞僩儕僾僔儞壛悈暘夝暔
嘇兝-僔僩僗僥儘乕儖
嘊IIAEK乮兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼棃偺儁僾僠僪乯
昞1丂幚尡怘偺慻惉乮g/300 g乯
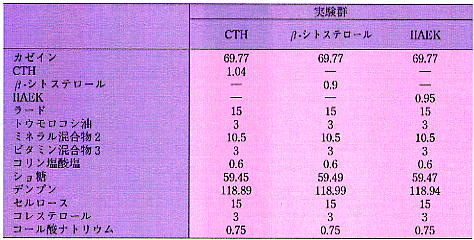 1. CTH偼僇僛僀儞僩儕僾僔儞壛悈暘夝暔傪偄偆
2. AIN-93G 儈僱儔儖崿崌暔. 怘塧偼丄師偺傛偆側儈僱儔儖傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
丂Ca:5000,P:1561,K:3600,S:300,Na:1019,Cl:1571,Mg:507,Fe:35,Zn:30,Mn:10,
丂Cu:6,I:0.2,Mo:0.15,Se:0.15,Si:5,Cr:1.0,F:1.0,Ni:0.5,B:0.5,Li:0.1,V:0.1.
3. AIN-76 價僞儈儞崿崌暔. 怘塧偼丄師偺傛偆側價僞儈儞傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
丂all-trans-retinyl palminate:4000IU,cholecalciferol:1000IU,
丂all-rac-兛-tocopheryl acetate:75IU,phylloquinone:0.75,
丂thiamine-HCl:6.36,rivoflavin:6.0,pyridoxin-HCl:7.29,nicotinic acid:30,
丂Ca-pantothnate:16.31,folic acid:2.0,cyanocobalamin:0.025,biotin:0.2.
幚尡2
丂摦暔幚尡乮儅僂僗乯偵傛傞兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼
棃偺儁僾僠僪乮GLDIQK丄ALPMH乯偺僐儗僗僥儘乕儖戙幱偵懳偡傞塭嬁偺専摙
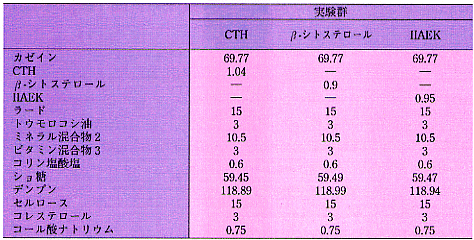
丂ICR宯梇儅僂僗乮弶懱廳栺20g乯傪梡偄偨丅20亾僇僛僀儞偵1亾偺僐儗僗僥儘
乕儖傪揧壛偟偨帞椏偵丄僇僛僀儞僩儕僾僔儞壛悈暘夝暔乮CTH乯丄兝-儔僋僩
僌儘僽儕儞僩儕僾僔儞壛悈暘夝暔乮LTH乯丄GLDIQK偍傛傃ALPMH傪偦傟偧傟0.3
亾揧壛偟偨怘塧傪4擔娫愛庢偝偣偨丅怘塧慻惉偼丄昞2偵帵偟偨丅4擔栚偵儔僢
僩傪怱憻嵦寣偵傛傝偲嶦偟偨丅奺孮偼偦傟偧傟6旵偲偟偨丅懱廳偼2擔枅偵24
帪娫偺怘塧愛庢検偼丄4擔栚偵應掕偟丄傆傫偺嵦庢偼2擔栚偐傜4擔栚偵峴偭偨丅
4擔栚偵儔僢僩傪僄乕僥儖杻悓壓偱怱憻嵦寣偵傛傝偲嶦偟丄娞憻傪揈弌偟偨丅
寣惔偼墦怱暘棧乮3000夞揮丄15暘乯偵傛傝挷惢偟丄暘愅傑偱搥寢曐懚偟偨丅
寣惔帀幙偼峺慺朄偱應掕偟偨丅娞憻偼昞柺偵晅偄偨寣塼側偳傪傛偔傆偒庢傝丄
0.9亾NaCl梟塼偱愻忩屻丄幖廳検傪應掕偟丄暘愅傑偱搥寢曐懚偟偨丅娞憻帀幙
偺拪弌丄惛惢偼丄Folch傜偺曽朄乮9乯偵廬偭偰峴偄丄峺慺朄偱應掕偟偨丅傆
傫拞僗僥儘僀僪攔煏検偼婛曬乮10乯偵廬偭偰専摙偟偨丅
幚尡孮偼壓婰偺傛偆側傕偺偱偁傞丅
嘆CTH丗僇僛僀儞僩儕僾僔儞壛悈暘夝暔
嘇LTH丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔
嘊GLDIQK丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼棃偺儁僾僠僪
嘋ALPMH丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼棃偺儁僾僠僪
[摑寁暘愅]
幚尡1丄2偺幚尡寢壥偺摑寁揑暘愅偵偼丄Duncan's Multiple Range Test傪梡
偄偨丅
昞2丂幚尡怘偺慻惉乮g/300 g乯
1. CTH偼僇僛僀儞僩儕僾僔儞壛悈暘夝暔傪偄偆
2. AIN-93G 儈僱儔儖崿崌暔. 怘塧偼丄師偺傛偆側儈僱儔儖傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
丂Ca:5000,P:1561,K:3600,S:300,Na:1019,Cl:1571,Mg:507,Fe:35,Zn:30,Mn:10,
丂Cu:6,I:0.2,Mo:0.15,Se:0.15,Si:5,Cr:1.0,F:1.0,Ni:0.5,B:0.5,Li:0.1,V:0.1.
3. AIN-76 價僞儈儞崿崌暔. 怘塧偼丄師偺傛偆側價僞儈儞傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
丂all-trans-retinyl palminate:4000IU,cholecalciferol:1000IU,
丂all-rac-兛-tocopheryl acetate:75IU,phylloquinone:0.75,
丂thiamine-HCl:6.36,rivoflavin:6.0,pyridoxin-HCl:7.29,nicotinic acid:30,
丂Ca-pantothnate:16.31,folic acid:2.0,cyanocobalamin:0.025,biotin:0.2.
幚尡2
丂摦暔幚尡乮儅僂僗乯偵傛傞兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼
棃偺儁僾僠僪乮GLDIQK丄ALPMH乯偺僐儗僗僥儘乕儖戙幱偵懳偡傞塭嬁偺専摙
丂ICR宯梇儅僂僗乮弶懱廳栺20g乯傪梡偄偨丅20亾僇僛僀儞偵1亾偺僐儗僗僥儘
乕儖傪揧壛偟偨帞椏偵丄僇僛僀儞僩儕僾僔儞壛悈暘夝暔乮CTH乯丄兝-儔僋僩
僌儘僽儕儞僩儕僾僔儞壛悈暘夝暔乮LTH乯丄GLDIQK偍傛傃ALPMH傪偦傟偧傟0.3
亾揧壛偟偨怘塧傪4擔娫愛庢偝偣偨丅怘塧慻惉偼丄昞2偵帵偟偨丅4擔栚偵儔僢
僩傪怱憻嵦寣偵傛傝偲嶦偟偨丅奺孮偼偦傟偧傟6旵偲偟偨丅懱廳偼2擔枅偵24
帪娫偺怘塧愛庢検偼丄4擔栚偵應掕偟丄傆傫偺嵦庢偼2擔栚偐傜4擔栚偵峴偭偨丅
4擔栚偵儔僢僩傪僄乕僥儖杻悓壓偱怱憻嵦寣偵傛傝偲嶦偟丄娞憻傪揈弌偟偨丅
寣惔偼墦怱暘棧乮3000夞揮丄15暘乯偵傛傝挷惢偟丄暘愅傑偱搥寢曐懚偟偨丅
寣惔帀幙偼峺慺朄偱應掕偟偨丅娞憻偼昞柺偵晅偄偨寣塼側偳傪傛偔傆偒庢傝丄
0.9亾NaCl梟塼偱愻忩屻丄幖廳検傪應掕偟丄暘愅傑偱搥寢曐懚偟偨丅娞憻帀幙
偺拪弌丄惛惢偼丄Folch傜偺曽朄乮9乯偵廬偭偰峴偄丄峺慺朄偱應掕偟偨丅傆
傫拞僗僥儘僀僪攔煏検偼婛曬乮10乯偵廬偭偰専摙偟偨丅
幚尡孮偼壓婰偺傛偆側傕偺偱偁傞丅
嘆CTH丗僇僛僀儞僩儕僾僔儞壛悈暘夝暔
嘇LTH丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔
嘊GLDIQK丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼棃偺儁僾僠僪
嘋ALPMH丗兝-儔僋僩僌儘僽儕儞偺僩儕僾僔儞壛悈暘夝暔桼棃偺儁僾僠僪
[摑寁暘愅]
幚尡1丄2偺幚尡寢壥偺摑寁揑暘愅偵偼丄Duncan's Multiple Range Test傪梡
偄偨丅
昞2丂幚尡怘偺慻惉乮g/300 g乯
 1. CTH偼僇僛僀儞僩儕僾僔儞壛悈暘夝暔傪丄LTH偼兝-儔僋僩僌儘僽儕儞偺僩
儕僾僔儞壛悈暘夝暔傪偦傟偧傟偄偆
2. AIN-93G 儈僱儔儖崿崌暔. 怘塧偼丄師偺傛偆側儈僱儔儖傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
Ca:5000,P:1561,K:3600,S:300,Na:1019,Cl:1571,Mg:507,Fe:35,Zn:30,Mn:10,
Cu:6,I:0.2,Mo:0.15,Se:0.15,Si:5,Cr:1.0,F:1.0,Ni:0.5,B:0.5,Li:0.1,V:0.1.
3. AIN-76 價僞儈儞崿崌暔. 怘塧偼丄師偺傛偆側價僞儈儞傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
all-trans-retinyl palminate:4000IU,cholecalciferol:1000IU,
all-rac-兛-tocopheryl acetate:75IU,phylloquinone:0.75,
thiamine-HCl:6.36,rivoflavin:6.0,pyridoxin-HCl:7.29,nicotinic acid:30,
Ca-pantothnate:16.31,folic acid:2.0,cyanocobalamin:0.025,biotin:0.2.
1. CTH偼僇僛僀儞僩儕僾僔儞壛悈暘夝暔傪丄LTH偼兝-儔僋僩僌儘僽儕儞偺僩
儕僾僔儞壛悈暘夝暔傪偦傟偧傟偄偆
2. AIN-93G 儈僱儔儖崿崌暔. 怘塧偼丄師偺傛偆側儈僱儔儖傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
Ca:5000,P:1561,K:3600,S:300,Na:1019,Cl:1571,Mg:507,Fe:35,Zn:30,Mn:10,
Cu:6,I:0.2,Mo:0.15,Se:0.15,Si:5,Cr:1.0,F:1.0,Ni:0.5,B:0.5,Li:0.1,V:0.1.
3. AIN-76 價僞儈儞崿崌暔. 怘塧偼丄師偺傛偆側價僞儈儞傪娷傫偱偄傞
乮mg/kg 怘塧乯丗
all-trans-retinyl palminate:4000IU,cholecalciferol:1000IU,
all-rac-兛-tocopheryl acetate:75IU,phylloquinone:0.75,
thiamine-HCl:6.36,rivoflavin:6.0,pyridoxin-HCl:7.29,nicotinic acid:30,
Ca-pantothnate:16.31,folic acid:2.0,cyanocobalamin:0.025,biotin:0.2.
尋媶寢壥
幚尡 1 乮摦暔幚尡乯乮昞 3 丄 4 丄 5 乯
丂懱廳憹壛検偵偍偄偰丄俠俿俫孮偲斾妑偟偰慡孮偱桳堄側嵎偼尒傜傟側偐偭偨丅
怘塧愛庢検偼丄孮娫偱曄壔偑側偐偭偨丅娞憻廳検偵偍偄偰丄CTH孮偲斾妑偟偰丄
俬俬俙俤俲孮偱桳堄偵忋徃偟偨丅寣惔帀幙暘愅偵偍偄偰丄憤僐儗僗僥儘乕儖抣
偼丄CTH孮偲斾妑偟偰IIAEK孮偱桳堄偵掅壓偟偨偑丄兝-僔僩僗僥儘乕儖孮偱偼
桳堄側掅壓偼惗偠側偐偭偨丅HDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑偟偰IIAEK
孮偵偍偄偰忋徃孹岦偑尒傜傟偨丅LDL+VLDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑
偟偰IIAEK孮偱桳堄偵掅壓偟偨丅憤僐儗僗僥儘乕儖偵偟傔傞HDL-僐儗僗僥儘乕
儖偺妱崌乮摦柆峝壔巜悢乯傪嶼弌偟偨偲偙傠丄CTH孮偲斾妑偟偰丄IIAEK孮偱桳
堄偵忋徃偟偨乮昞3乯丅
丂娞憻帀幙暘愅偵偍偄偰丄娞憻1僌儔儉摉偨傝偱偼丄憤帀幙抣偼丄CTH孮偲斾妑
偟偰兝-僔僩僗僥儘乕儖孮偱桳堄側掅壓偑尒傜傟丄IIAEK孮偱掅壓孹岦偑尒傜傟
偨丅僐儗僗僥儘乕儖抣偼丄IIAEK孮偱CTH孮偲斾傋偰桳堄側掅壓偑尒傜傟偨偑丄
兝-僔僩僗僥儘乕儖孮偱偼桳堄側嵎偼尒傜傟側偐偭偨丅100僌儔儉懱廳摉偨傝偵
姺嶼偡傞偲丄憤帀幙抣媦偵偍偄偰丄CTH孮偲斾妑偡傞偲兝-僔僩僗僥儘乕儖孮偼
桳堄偵掅壓偟偨偑丄IIAEK孮偱偼桳堄側嵎偼尒傜傟側偐偭偨丅僐儗僗僥儘乕儖
抣偵偍偄偰丄孮娫偱桳堄嵎偼尒傜傟側偐偭偨乮昞4乯丅
丂傆傫拞僗僥儘僀僪暘愅偵偍偄偰丄巁惈僗僥儘僀僪攔煏検偼丄CTH孮偲斾妑偟
偰丄IIAEK孮偱桳堄偵忋徃偟丄兝-僔僩僗僥儘乕儖孮偱偼掅壓孹岦傪帵偟偨丅傆
傫拞偺僐儗僗僥儘乕儖偺攔煏検偼丄CTH孮偲斾妑偟偰丄兝-僔僩僗僥儘乕儖孮丄
IIAEK孮偱桳堄偵忋徃偟偨丅拞惈僗僥儘僀僪攔煏検偵偍偄偰傕丄CTH孮偲斾妑偟
偰丄兝-僔僩僗僥儘乕儖孮丄IIAEK孮偱桳堄偵忋徃偟偨丅廬偭偰丄憤僗僥儘僀僪
攔煏検偵偍偄偰丄CTH孮偲斾妑偟偰丄兝-僔僩僗僥儘乕儖孮丄IIAEK孮偱桳堄偵
忋徃偟偨丅乮昞5乯
昞3丂儅僂僗偵偍偗傞CTH丄兝-僔僩僗僥儘乕儖丄IIAEK偺懱廳憹壛検丄
娞憻廳検丄怘塧愛庢検丄寣惔帀幙偵懳偡傞塭嬁 1 , 2 , 3
 1. 愛庢婜娫偼 4 擔娫. 丂丂丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎. 4. LDL+VLDL-僐儗僗僥儘乕儖亖寣惔憤僐儗僗僥儘乕儖 - HDL僐儗僗僥儘乕儖
昞4丂儅僂僗偵偍偗傞CTH丄兝-僔僩僗僥儘乕儖丄IIAEK偺娞憻帀幙偵懳偡傞塭嬁 1 , 2 , 3
1. 愛庢婜娫偼 4 擔娫. 丂丂丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎. 4. LDL+VLDL-僐儗僗僥儘乕儖亖寣惔憤僐儗僗僥儘乕儖 - HDL僐儗僗僥儘乕儖
昞4丂儅僂僗偵偍偗傞CTH丄兝-僔僩僗僥儘乕儖丄IIAEK偺娞憻帀幙偵懳偡傞塭嬁 1 , 2 , 3
 1. 愛庢婜娫偼 4 擔娫.丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
昞5丂儅僂僗偵偍偗傞CTH丄兝-僔僩僗僥儘乕儖丄IIAEK偺傆傫拞帀幙偵懳偡傞塭嬁 1 , 2 , 3
1. 愛庢婜娫偼 4 擔娫.丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
昞5丂儅僂僗偵偍偗傞CTH丄兝-僔僩僗僥儘乕儖丄IIAEK偺傆傫拞帀幙偵懳偡傞塭嬁 1 , 2 , 3
 1. 愛庢婜娫偼 4 擔娫.丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
幚尡 2 乮摦暔幚尡乯乮昞 6 丄 7 丄 8 乯
丂懱廳憹壛検丄怘塧愛庢検偵偍偄偰丄孮娫偱桳堄嵎偼尒傜傟側偐偭偨丅娞憻廳
検偵偍偄偰丄CTH孮偲斾妑偟偰丄GLDIQK孮乮埲壓G孮偲偄偆丅乯丄ALPMH孮乮埲
壓A孮偲偄偆丅乯偱桳堄偵忋徃偟偨丅寣惔帀幙暘愅偵偍偄偰丄憤僐儗僗僥儘乕
儖抣偼丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄偵掅壓偟偨偑丄LTH孮偱偼桳堄側掅
壓偼惗偠側偐偭偨丅HDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑偟偰丄慡孮偱桳堄
側嵎偼尒傜傟側偐偭偨丅LDL+VLDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑偟偰丄G
孮丄A孮偱桳堄偵掅壓偟偨丅憤僐儗僗僥儘乕儖偵偟傔傞HDL-僐儗僗僥儘乕儖偺
妱崌乮摦柆峝壔巜悢乯傪嶼弌偟偨偲偙傠丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄
偵忋徃偟偨丅乮昞6乯
昞俇丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮偺懱廳憹壛検丄娞憻廳検丄
怘塧愛庢検丄寣惔帀幙偵懳偡傞塭嬁1,2,3
1. 愛庢婜娫偼 4 擔娫.丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
幚尡 2 乮摦暔幚尡乯乮昞 6 丄 7 丄 8 乯
丂懱廳憹壛検丄怘塧愛庢検偵偍偄偰丄孮娫偱桳堄嵎偼尒傜傟側偐偭偨丅娞憻廳
検偵偍偄偰丄CTH孮偲斾妑偟偰丄GLDIQK孮乮埲壓G孮偲偄偆丅乯丄ALPMH孮乮埲
壓A孮偲偄偆丅乯偱桳堄偵忋徃偟偨丅寣惔帀幙暘愅偵偍偄偰丄憤僐儗僗僥儘乕
儖抣偼丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄偵掅壓偟偨偑丄LTH孮偱偼桳堄側掅
壓偼惗偠側偐偭偨丅HDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑偟偰丄慡孮偱桳堄
側嵎偼尒傜傟側偐偭偨丅LDL+VLDL-僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑偟偰丄G
孮丄A孮偱桳堄偵掅壓偟偨丅憤僐儗僗僥儘乕儖偵偟傔傞HDL-僐儗僗僥儘乕儖偺
妱崌乮摦柆峝壔巜悢乯傪嶼弌偟偨偲偙傠丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄
偵忋徃偟偨丅乮昞6乯
昞俇丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮偺懱廳憹壛検丄娞憻廳検丄
怘塧愛庢検丄寣惔帀幙偵懳偡傞塭嬁1,2,3
 1. 愛庢婜娫偼 4 擔娫. 丂丂丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎. 4. LDL+VLDL-僐儗僗僥儘乕儖亖寣惔憤僐儗僗僥儘乕儖 - HDL僐儗僗僥儘乕儖
丂娞憻帀幙暘愅偵偍偄偰丄娞憻1僌儔儉摉偨傝偱偼丄憤帀幙抣偼丄CTH孮偲斾妑
偟偰丄A孮偼Student's t-test偵偍偄偰桳堄偵掅壓偟偨丅拞惈帀幙偍傛傃儕儞
帀幙偼丄CTH孮偲斾妑偟偰丄慡孮偱桳堄側嵎偼尒傜傟側偐偭偨丅僐儗僗僥儘乕
儖抣偼丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄偵掅壓偟偨丅100僌儔儉懱廳摉偨傝
偵姺嶼偡傞偲丄憤帀幙抣偍傛傃拞惈帀幙抣偱偼丄CTH孮偲斾妑偟偰丄慡孮偱桳
堄側嵎偼尒傜傟側偐偭偨丅儕儞帀幙抣偼丄CTH孮偲斾妑偟偰丄LTH孮偲G孮偱忋
徃孹岦傪帵偟丄A孮偱桳堄偵忋徃偟偨丅傑偨僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑
偟偰丄LTH孮丄A孮偱掅壓孹岦傪帵偟丄G孮偱桳堄偵掅壓偟偨丅乮昞7乯
丂傆傫拞僗僥儘僀僪暘愅偵偍偄偰丄巁惈僗僥儘僀僪攔煏検偼丄CTH孮偲斾妑偟
偰丄G孮丄A孮偱桳堄偵忋徃偟偨丅傆傫拞偺僐儗僗僥儘乕儖偺攔煏検偼丄CTH孮
偲斾妑偟偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄A孮偱桳堄偵忋徃偟偨丅拞惈僗僥
儘僀僪攔煏検偵偍偄偰傕丄CTH孮偲斾妑偟偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄
A孮偱桳堄偵忋徃偟偨丅廬偭偰丄憤僗僥儘僀僪攔煏検偵偍偄偰丄CTH孮偲斾妑偟
偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄A孮偱桳堄偵忋徃偟偨乮昞8乯
昞7丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮偺娞憻帀幙偵懳偡傞塭嬁 1 , 2 , 3
1. 愛庢婜娫偼 4 擔娫. 丂丂丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎. 4. LDL+VLDL-僐儗僗僥儘乕儖亖寣惔憤僐儗僗僥儘乕儖 - HDL僐儗僗僥儘乕儖
丂娞憻帀幙暘愅偵偍偄偰丄娞憻1僌儔儉摉偨傝偱偼丄憤帀幙抣偼丄CTH孮偲斾妑
偟偰丄A孮偼Student's t-test偵偍偄偰桳堄偵掅壓偟偨丅拞惈帀幙偍傛傃儕儞
帀幙偼丄CTH孮偲斾妑偟偰丄慡孮偱桳堄側嵎偼尒傜傟側偐偭偨丅僐儗僗僥儘乕
儖抣偼丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄偵掅壓偟偨丅100僌儔儉懱廳摉偨傝
偵姺嶼偡傞偲丄憤帀幙抣偍傛傃拞惈帀幙抣偱偼丄CTH孮偲斾妑偟偰丄慡孮偱桳
堄側嵎偼尒傜傟側偐偭偨丅儕儞帀幙抣偼丄CTH孮偲斾妑偟偰丄LTH孮偲G孮偱忋
徃孹岦傪帵偟丄A孮偱桳堄偵忋徃偟偨丅傑偨僐儗僗僥儘乕儖抣偼丄CTH孮偲斾妑
偟偰丄LTH孮丄A孮偱掅壓孹岦傪帵偟丄G孮偱桳堄偵掅壓偟偨丅乮昞7乯
丂傆傫拞僗僥儘僀僪暘愅偵偍偄偰丄巁惈僗僥儘僀僪攔煏検偼丄CTH孮偲斾妑偟
偰丄G孮丄A孮偱桳堄偵忋徃偟偨丅傆傫拞偺僐儗僗僥儘乕儖偺攔煏検偼丄CTH孮
偲斾妑偟偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄A孮偱桳堄偵忋徃偟偨丅拞惈僗僥
儘僀僪攔煏検偵偍偄偰傕丄CTH孮偲斾妑偟偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄
A孮偱桳堄偵忋徃偟偨丅廬偭偰丄憤僗僥儘僀僪攔煏検偵偍偄偰丄CTH孮偲斾妑偟
偰丄LTH孮偱忋徃孹岦傪帵偟丄G孮丄A孮偱桳堄偵忋徃偟偨乮昞8乯
昞7丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮偺娞憻帀幙偵懳偡傞塭嬁 1 , 2 , 3
 1. 愛庢婜娫偼 4 擔娫.丂丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
昞8丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮 偺傆傫拞帀幙偵懳偡傞塭嬁 1 , 2 , 3
1. 愛庢婜娫偼 4 擔娫.丂丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
昞8丂儅僂僗偵偍偗傞CTH丄LTH丄G孮丄A孮 偺傆傫拞帀幙偵懳偡傞塭嬁 1 , 2 , 3
 1. 愛庢婜娫偼 4 擔娫.丂丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
1. 愛庢婜娫偼 4 擔娫.丂丂丂2. 奺孮 6 旵偺暯嬒抣亇昗弨曃嵎.丂丂丂3. 堎側偭偨暥帤娫偱桳堄嵎偁傝乮p<0.05乯.
尋媶峫嶡
丂偙傟傑偱偺尋媶偐傜丄戝摛僞儞僷僋幙偺寣惔僐儗僗僥儘乕儖掅壓嶌梡偵偼丄
慳悈惈偺儁僾僠僪偑娭梌偡傞偙偲偑壖愢偲偟偰峫偊傜傟偰偒偨乮11丄12乯丅
偟偐偟丄in vivo偱傕妶惈傪敪尰偡傞戝摛僞儞僷僋幙桼棃偺寣惔僐儗僗僥儘
乕儖掅尭壔儁僾僠僪偼枹偩偵枹敪尒偱偁傞丅傢傟傢傟偼悽奅偱弶傔偰丄兝-儔
僋僩僌儘僽儕儞桼棃偺怴偟偄寣惔僐儗僗僥儘乕儖掅尭壔儁僾僠僪傪3庬椶乮II
AEK丄GLDIQK丄ALPMH乯敪尒偟偨丅
丂崱夞偺尋媶偵傛傝丄儅僂僗偵偍偄偰丄帞椏偵兝-儔僋僩僌儘僽儕儞桼棃儁僾
僠僪乮IIAEK乯傪0.3亾偲偄偆嬌旝検揧壛偡傞偙偲偵傛傝丄寣惔僐儗僗僥儘乕
儖偼寑揑偵掅壓偟偨丅偟偐傕IIAEK偼丄堛栻昳偱偁傞兝-僔僩僗僥儘乕儖傛傝
傕嫮椡側寣惔僐儗僗僥儘乕儖掅壓嶌梡傪敪尰偟偨丅崱夞丄弶傔偰儅僂僗傪梡
偄偨抁婜昡壙宯偑峔抸偝傟偨偙偲偐傜丄崱屻偼丄嬌旝検偱崅壙側崌惉儁僾僠
僪偺昡壙偑壜擻偲側傠偆丅傑偨丄宱岥搳梌偲斾傋偰憖嶌偑梕堈偱偁傞偙偲偐
傜丄崱夞偺幚尡宯傕嬌傔偰桳梡偱偁傠偆丅IIAEK愛庢帪偵偍偄偰丄傆傫拞傊偺
僗僥儘僀僪攔煏検傪應掕偟偨偑丄懳徠孮乮CTH孮乯偲斾妑偟偰丄桳堄側傆傫拞
傊偺僗僥儘僀僪攔煏検偺憹壛偑娤嶡偝傟偨丅IIAEK偼丄彫挵偵偍偗傞僐儗僗僥
儘乕儖偺媧廂傪慾奞偟偰偄傞壜擻惈偑帵嵈偝傟偨丅
丂偝傜偵丄兝-儔僋僩僌儘僽儕儞桼棃儁僾僠僪偱偁傞G孮偍傛傃A孮傪怘塧偲偟
偰儅僂僗偵梌偊偨帪偺塭嬁傪専摙偟偨丅G孮傗A孮偼丄懳徠孮乮CTH孮乯偵斾傋
偰寣惔僐儗僗僥儘乕儖儗儀儖傪寑揑偵掅壓偝偣偨丅娞憻僐儗僗僥儘乕儖儗儀
儖偼丄娞憻1僌儔儉摉偨傝偵偍偄偰丄CTH孮偲斾妑偟偰丄G孮丄A孮偱桳堄側掅
壓傪帵偟丄G孮偱偼100僌儔儉懱廳摉偨傝偵姺嶼偟偨偁偲偱傕丄CTH孮偲斾妑偟
偰桳堄側掅壓偑娤嶡偝傟偨丅傑偨G孮丄A孮偱偼丄CTH孮偲斾妑偟偰丄傆傫拞偺
巁惈僗僥儘僀僪攔煏偲拞惈僗僥儘僀僪攔煏偺偄偢傟傕桳堄偵忋徃偟偨丅偙偺
偙偲偐傜G孮丄A孮偱偼丄彫挵偵偍偗傞僐儗僗僥儘乕儖偺媧廂傪慾奞偟偰偄傞
壜擻惈偑帵嵈偝傟偨丅崱屻偼丄偙傟傜偺儁僾僠僪偑丄偳偺傛偆側婡峔偱傆傫
拞傊偺僗僥儘僀僪攔煏憹壛傪桿摫偡傞偺偐傪夝柧偟偨偄偲峫偊傞丅傑偨IIAEK丄
GLDIQK傗ALPMH偑丄偳偺掱搙偳偺傛偆側宍懺偱媧廂偝傟丄媧廂屻偵娞憻側偳偺
僐儗僗僥儘乕儖戙幱娭楢堚揱巕敪尰偵懳偟偰丄偳偺傛偆偵塭嬁偡傞偺偐傪専摙
偡傞偺偼戝曄嫽枴怺偄偲峫偊傞丅
丂廬棃丄僞儞僷僋幙傗傾儈僲巁偺僐儗僗僥儘乕儖戙幱偵懳偡傞塭嬁偼丄儔僢僩
側偳偵丄5亾埲忋偺儗儀儖偱怘塧偵揧壛偟偨応崌偵偍偄偰娤嶡偝傟偰偄傞応崌
偑懡偄丅偟偐偟丄崱夞偺尋媶偱敪尒偟偨3庬椶偺怴偟偄寣惔僐儗僗僥儘乕儖掅
尭壔儁僾僠僪偼丄傢偢偐怘塧偵0.3亾揧壛偱嬌傔偰嫮椡側岠壥傪敪婗偟偨丅偙
偺偙偲偐傜丄廬棃偺僞儞僷僋幙帺恎傗傾儈僲巁扨撈偺愛庢偵傛傞僐儗僗僥儘乕
儖戙幱偵懳偡傞塭嬁偐傜峫偊傜傟偰偒偨奣擮乮13乣16乯偱偼丄偙傟傜偺儁僾僠
僪偺岠壥偼愢柧晄壜擻偱偁傝丄儁僾僠僪偺僐儗僗僥儘乕儖戙幱偵懳偡傞塭嬁傪
愢柧偡傞偨傔偺怴偟偄奣擮偺峔抸偑昁梫偱偁傞丅偦偺堄枴偱傕偙傟傜3庬椶偺
寣惔僐儗僗僥儘乕儖掅尭壔儁僾僠僪偺敪尒偼拲栚偵抣偡傞偲峫偊傜傟傞丅
丂埲忋偺傛偆側尋媶偐傜丄兝-儔僋僩僌儘僽儕儞桼棃偺儁僾僠僪偱偁傞IIAEK埲
奜偵丄GLDIQK偍傛傃ALPMH偑丄寣惔僐儗僗僥儘乕儖掅壓嶌梡傪敪尰偡傞偙偲傪
悽奅偵愭嬱偗偰弶傔偰敪尒偟偨丅偙傟傜偺儁僾僠僪偺僐儗僗僥儘乕儖戙幱夵慞
嶌梡偼丄偙傟傑偱偵曬崘偑側偔丄怴偨側抺嶻帒尮偺桳梡惈偺敪尒偲偟偰拲栚偵
抣偡傞丅偙傟傜偺偙偲偼丄抺嶻暔偺廀梫奐敪丄廀梫憹恑偵柧傜偐偵寢傃偮偔傕
偺偱偁傞丅
嶲峫榑暥
1丏Carrol, K. K. and Hamilton, R. M. G. 乮1975乯
丂Effects of dietary protein and carbohydrate on plasma cholesterol
丂levels in relation to atherosclerosis. J.Food Sci., 40, 18-23.
2丏Nagaoka, S., Kanamaru, Y. and Kuzuya,Y. 乮1991乯
丂Effects of whey protein and casein on the plasma and liver lipids
丂in rats. Agric.Biol.Chem., 55, 813-818.
3丏Nagaoka, S., Kanamaru, Y., Kuzuya, Y., Kojima, T and Kuwata, T.
丂乮1992乯 Comparative studies on the serum cholesterol lowering
丂action of whey protein and soybean protein in rats. Biosci.Biotech.
丂Biochem., 56, 1484-1485.
4丏挿壀丂棙丄嬥娵媊宧丄妺扟懽梇丗擕惔僞儞僷僋幙偺寣惔僐儗僗僥儘乕儖
丂掅壓嶌梡丄嶳撪朚抝丄崱懞宱柧丄庣揷揘楴丂愑擟曇廤"媿擕惉暘偺摿惈偲
丂寬峃"搶嫗丗岝惗娰丄57-84 乮1993乯
5丏挿壀丂棙丗怘昳惉暘偍傛傃惗懱堎暔偵傛傞僐儗僗僥儘乕儖戙幱惂屼偵
丂娭偡傞尋媶乮擔杮塰梴怘椘妛夛彠椼徿庴徿憤愢乯擔杮塰梴怘椘妛夛帍丄
丂49, 303-313 乮1996乯
6丏Nagata, Y., Ishiwaki, N and Sugano, M. 乮1982乯
丂Studies on the mechanisms of antihypercholesterolemic action of
丂soy protein and soy protein-type amino mixtures in relation to
丂the casein counterparts in rats. J.Nutr., 112, 1614-1625.
7丏Nagaoka, S., Awano, T., Nagata, N., Masaoka, M., Hori, G. and
丂 Hashimoto, K. 乮1997乯 Serum cholesterol reduction and cholesterol
丂absorption inhibition in CaCo-2 cells by a soyprotein peptic
丂hydrolyzate. Biosci.Biotech.Biochem., 61, 354-356.
8丏Nagaoka, S., Futamura, Y., Miwa, K., Awano, T., Yamauchi,
丂 K., Kanamaru, Y., Kojima, T. and Kuwata, T.: Identification of
丂novel hypocholesterolemic peptides derived from bovine milk
丂兝-lactoglobulin. Biochem.Biophys.Res.Commun., 281, 11-17 乮2001乯
9丏Folch, J., Lees, M. and Sloane-Stanley, G. H. 乮1957乯
丂A simple method for the isolation and purification of total
丂lipids from animal tissues. J. Biol. Chem., 226, 497-509.
10丏Nagaoka, S., Miwa, K., Eto, M., Kuzuya, Y., Hori, G.
丂 and Yamamoto, K. 乮1999乯 Soyprotein peptic hydrolyzate with bound
丂 phospholipids decrease micellar solubility and cholesterol
丂 absorption in rats and Caco-2 cells. J. Nutr., 129, 1725-1730.
11丏Iwami, K., Sakakibara, K. and Ibuki, F. 乮1986乯
Involvement of post- digestion 'hydrophobic' peptides in plasma
cholesterol-lowering effect of dietary plant proteins.
Agric.Biol.Chem., 50, 1217-1222.
12丏Sugano, M., Goto, S., Yamada, Y., Yoshida, K., Hashimoto,
Y., Matsuo, T. and Kimoto, M. 乮1990乯 Cholesterol-lowering
activity of various undigested fractions of soybean protein
in rats. J. Nutr., 120, 977-985.
13丏Huff, M.W. and Carroll, K.K. 乮1980乯
Effects of dietary proteins and amino acid mixtures on plasma
cholesterol levels in rabbits. J. Nutr., 110, 1676-1685.
14丏Jacques, H., Deshaies, Y. and Savoie, L. 乮1986乯
Relationship between dietary proteins, their in vitro
digestion products, and serum cholesterol in rats.
Atherosclerosis, 61, 89-98.
15丏Nagata, Y., Ishiwaki, N and Sugano, M. 乮1982乯
Studies on the mechanisms of antihypercholesterolemic action
of soy protein and soy protein-type amino mixtures in relation
to the casein counterparts in rats. J. Nutr., 112, 1614-1625.
16丏Sugiyama, K., Ohkawa, S. and Muramatsu, K. 乮1986乯
Relationship between amino acid composition of diet and plasma
cholesterol level in growing rats fed a high cholesterol diet.
J.Nutr.Sci.Vitaminol., 32, 413-423.
尦偺儁乕僕偵栠傞