図1 オオシモフリエダシャクの明色型と暗色型
図1 オオシモフリエダシャクの明色型と暗色型
京都産業大学 工学部 生物工学科
教授 野村哲郎
黒毛和種は現在、約60万頭の繁殖雌牛が全国規模で飼養されており、和牛4 品種の中で、その重要性は頭数的、経済的に他の3品種を圧倒している。筆者 は黒毛和種集団の遺伝的多様性の経時的推移を調べるための研究を行ってきた。 本稿では、遺伝的多様性の育種的意義について解説した上で、黒毛和種集団に 関する分析結果を示し、本品種の抱える育種上の課題について述べる。
遺伝的多様性とは、特定の形質あるいは遺伝子座についての遺伝的変異のこ とである。ヒトのABO血液型では、これを決める遺伝子座について集団内に3 種の遺伝子があり、遺伝的変異が存在している。動物の育種において、選抜( 望ましい性質を持つ個体を親として利用するために選ぶこと)によって次世代 が遺伝的に改良されるためには、選抜の基準とした性質に遺伝的変異があり、 親として選んだ個体が望ましい遺伝子を持つことが前提となる 品種の持つ遺伝的多様性の大きさは、経済形質の遺伝的改良という比較的短 期間の育種成果を決める要因であるだけでなく、長期的な見地からも重要な意 義を持つ。この点は、自然集団における進化を例にして考えれば理解しやすい。 英国に生息するガの1種、オオシモフリエダシャクに認められた「工業暗化」 は、人類が自然選択を目撃できた数少ない例としてしばしば引用される現象で ある。このガには遺伝的に決定される「明色型」と「暗色型」と呼ばれる2つ のタイプがある(図1)。1850年当時は、このガがとまる木の幹は白っぽい
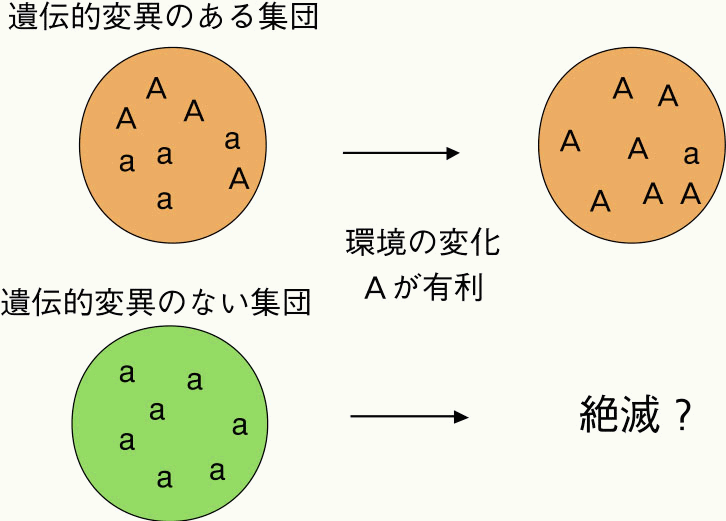
地衣類に覆われていたため、明色型は鳥からの捕食を逃れるための保護色とし て働いていた。逆に、暗色型は目立つために不利と考えられていた。ところが、 石炭の利用による大気汚染が進んだ50年後には、有利不利の関係が逆転した。 大気汚染により地衣類が脱落した木の幹は黒っぽいため、それまでは不利であ った暗色型が一転して有利になったのである。図2に示すように、マンチェス 図2 オオシモフリエダシャクの暗色型の割合の年次変化図ターでは1850年には暗色型の割合は1%以下であったが、工業化の進んだ5 0 年後には99%に増加していた。その後、大気汚染の人体への影響が指摘され、 汚染された環境が徐々に回復するにつれて、再び暗色型の割合が減少し、明色 型の割合が増加し始めている。もし、このガの集団に色彩に関して遺伝的多様 性がなければ(明色型しか存在しなければ)、このガは大気汚染という環境の 変化を乗り切ることができずに絶滅していたかもしれない(図3)。育種的な 見地から見ても、遺伝的多様性は、将来の育種目標の変更などに品種が迅速に 対応できるかどうかを決定する要因である。 図3 遺伝変異がある集団とない集団の比較
集団内の遺伝的多様性を測るための尺度にはいろいろなものがあるが、ここ ではもっとも簡便な遺伝分散の大きさを考える。ただし、他の尺度で表しても 本質的に同じ結論が導かれる。 いま、ある世代(t)での遺伝分散がVG,tである集団を考えよう。次の世代 (t+1)での遺伝分散は、前の世代より小さくなることが知られている。2つ の世代の遺伝分散の間には、図4に示したような関係が成り立つ。この関係か 図4 親世代と子世代での遺伝分散の関係
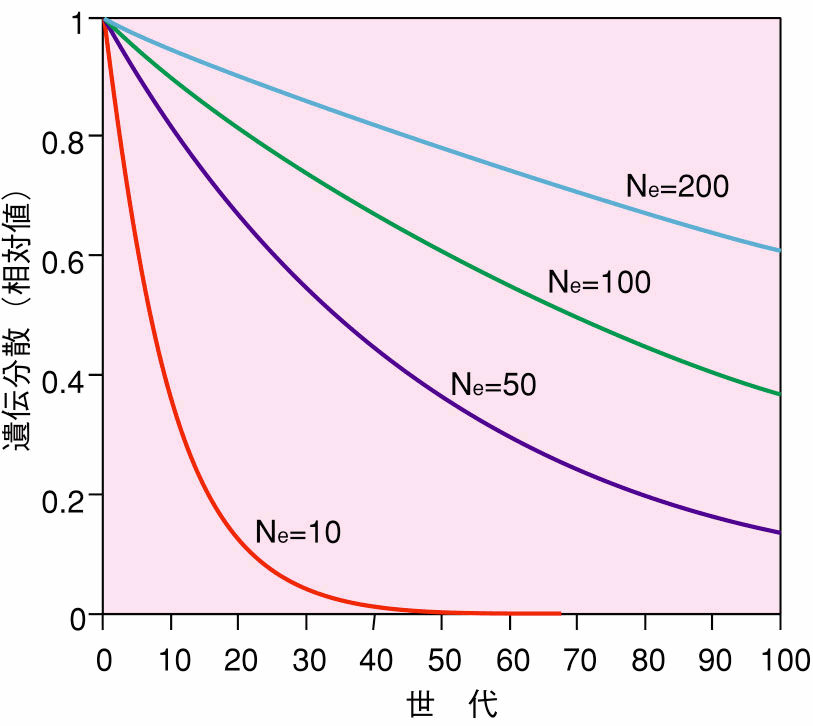
ら、世代交代に伴う遺伝分散の減少には、「集団の有効な大きさ」が関与して いることがわかる。集団の有効な大きさとは、簡単に言うと遺伝学的に有効な 個体数のことである。次の世代が雌雄1組から作られた場合と雌雄50組から作 られた場合では、後者のほうが遺伝的多様性に富んだ集団となることを考えれ ば、世代交代に伴う遺伝分散の減少に個体数(親の数)が関係することは直感 的に理解できるであろう。 図5には、いろいろな集団の有効な大きさの下での、遺伝分散(遺伝的多様 性)の世代の進行に伴う変化を示しておく。 図5 いろいろな集団の有効な大きさ(Ne)の下での 遺伝分散の世代に伴う変化
図4の関係式中の集団の有効な大きさとは、実際の牛を1頭、2頭というよう に数えたもの(見かけの個体数)とは異なる。以下では、見かけの個体数をN、 集団の有効な大きさをNeという記号で表して区別することにする。見かけの 個体数と集団の有効な大きさを結び付けるためには、いくつかの公式がある。 それらの公式を実際の家畜集団のように複雑な繁殖構造を持つ集団に当てはめ ることはできないが、集団の有効な大きさの性質を理解する上で重要と考えら れるので、図6に示しておく。 図6 集団の有効な大きさの代表的な公式
図6の(1)式は、雌雄同数のN個体がランダムにペアを作って、それぞれの ペアから子どもが生まれるような集団を考えたときの公式である。具体的にイ メージするために、ヒトの集団を考えてみよう。 個体数(人口)は毎世代おおよそ一定とすると、各ペア(夫婦)は、平均し て2人の子どもを残しているはずである。ただし、夫婦間には子どもの数にバ ラツキがあり、このバラツキを表したものが式の中のVkである。ヒトの多く の集団では、おおよそVk=2に近いことが知られている。これを(1)式に代 入するとNe=Nとなり、この場合には集団の有効な大きさは、見かけの個体数 に等しくなることがわかる。 黒毛和種集団では、周知のように、種雄牛の供用が人気のある少数の個体に 極端に集中している。従って、特定の親が極端に多くの子を残していることに なり、(1)式の中のVkは2よりもはるかに大きな値をとる。Vkが式の分母に 入っていることからわかるように、種雄牛の供用の偏りは、集団の有効な大き さを小さくするように働く。 図6の(2)式は、雄親と雌親の数が異なる集団の有効な大きさを計算する ための公式である。例えば、次のような2つの集団を考えてみよう。 集団1:雄親50頭、雌親50頭 (Nm=50、Nf=50) 集団2:雄親10頭、雌親90頭 (Nm=10、Nf=90) 2つの集団の見かけの個体数(N=Nm+Nf)は、ともに100となる。しかし、 (2)式から集団の有効な大きさを計算すると、 集団1:Ne=100 集団2:Ne=36 となり、雄親の数と雌親の数がアンバランスな集団では、集団の有効な大きさ が見かけの個体数よりもはるかに小さくなることがわかる。黒毛和種集団のよ うに、種雄牛の数が繁殖雌牛の数よりもはるかに少ないときにも同じことが起 こる。
図7に、1981年から2000年までの黒毛和種集団の有効な大きさの推定値を 示した(注)。全国で毎年、数万頭が新たに登録されている黒毛和種集団の有 効な大きさが、数10頭に過ぎないことに、少々驚かれた方も多いと思う。こ のような「見かけの個体数」との大きな隔たりは、すでに述べたように「種雄 牛の供用頻度に大きなバラツキがあること」と「種雄牛の数と繁殖雌牛の数に 大きな違いがあること」の2つの要因によるものである。 図7 黒毛和種集団の有効な大きさ(Ne)の推定値
年ごとの変化について見てみると、1990年までの集団の有効な大きさは変動 しているが、おおよそ30−40の値を保っていた。しかし1991年以降、集団の有 効な大きさは急激に減少する傾向にあった。特に1993年から1997年の間の平均 は、わずか17.1である。その後、1997年からは若干回復する傾向もうかがえる が、依然として30前後の値である。これらの値を実感するには、ヒトの集団を 考えればよい。すでに述べたように、ヒトの集団は集団の有効な大きさが見か けの個体数にかなり近いとされている。従って、有効な大きさが30前後という 集団では、おおよそ男女15人からなる集団と同じ速度で遺伝的多様性が失われ ていることを示している。集団の規模がこの程度であると、数世代のうちに、 どの男女ペアの間にも血縁関係が生じて近交度が速やかに上昇し、集団の遺伝 的多様性が急激に枯渇することは想像に難くない。 図8には、1986年から2000年の間の新規登録牛の父親として現れた種雄牛の 数、および種雄牛当たりの後代数の分散を示した。これらは、それぞれ図6の 式(1)と(2)の中のNmおよびVkに対応している。供用された種雄牛の数は、 1990年までは1,000頭以上を保っていたが、その後は減少する傾向にあり、19 97年以降は750頭を割っている。種雄牛当たりの後代数の分散については、調 査期間の後半から急激に増加している。 図8 各年度の種雄牛数と種雄牛当たりの後代数の分散
このような変化には、1991年に実施に踏み切られた牛肉輸入の自由化が大 きく関与している。すなわち、輸入自由化以降、国際競争下で高品質の牛肉生 産が求められ、それに伴い肉質(脂肪交雑)に優れた遺伝的能力(育種価)を 持つ種雄牛への供用の集中と、逆に肉質に比較的劣ると思われる種雄牛の淘汰 が、種雄牛数の減少と後代数の分散の顕著な増大をもたらしたものと考えられ る。 すでに述べたように、これらの変化はともに集団の有効な大きさを縮小する 方向に働くが、変化の相対的な大きさ(図8)から見て、種雄牛当たりの後代 数の分散の急激な増大が集団の有効な大きさの縮小の主因となったものと考え られる。 図9は、各調査年で後代数の多かった上位5頭の種雄牛の後代が、その年の 全登録牛に占める割合を示したものである。この図から、少数の種雄牛への供 用の集中の進展が明瞭に見てとれる。特に、牛肉の輸入自由化が実施された1 991年以降にその集中が顕著になり始め、2000年では全登録牛の半数以上が、 わずか5頭の種雄牛の後代となっている。 図9 各年において後代数が多かった上位5頭の種雄牛の 後代が全登録牛に占める割合
比較のために、1960年と1980年の黒毛和種集団の有効な大きさの推定値と、 諸外国の家畜品種の有効な大きさの推定値を表に示した。この表から1960年当 時の黒毛和種集団の有効な大きさは、他品種に比べて特異的に大きかったこと がわかる。これは、県を単位として育種が進められてきたことを反映して集団 が著しい分化構造を呈し、当時の品種内に多様な遺伝変異が含まれていたこと を示している。その後、1980年には有効な大きさは約120にまで縮小している が、この値は諸外国の家畜品種における推定値に比較的近い。 表 1960年および1980年の黒毛和種集団の有効な大きさと 諸外国の家畜集団の有効な大きさ
黒毛和種の1960年の推定値を除いて見ると、興味深い点は集団の有効な大 きさが、現実の品種の大きさにまったく依存しておらず、いずれの品種でも1 00前後の値を持つことである。このことは、育種に実際に関与する有効な種 雄の数がほぼ一定していることを意味している。現在の黒毛和種集団の有効な 大きさは、多くの品種に認められるスタンダードな値の1/3から1/5以下で ある。 さらに注意しておきたいことは、米国のヘレフォード種やホルスタイン種は、 世界各国に飼養されているこれらの品種から見れば、一地方品種である点であ る。これらの地方品種には、さらにそれらを包括する上位集団が存在し、地方 品種は上位集団から必要に応じて常に遺伝子の供給を受けることができる構造 を持っている。これに対して、黒毛和種は最上位のレベルで、集団の有効な大 きさの縮小と遺伝的多様性の低下が進行していることは十分に認識しておくべ きであろう。
和牛の育種において、遺伝的多様性の維持はしばしば問題にされてきた点で ある。品種が遺伝的構成の異なる分集団を内包していたことは、一面において 品種を斉一性の欠くものとしてきたことは事実である。しかし、品種内の遺伝 的多様性は、過去の改良過程において重要な働きを持ったと考えられる。例え ば、品種として成立した当時の黒毛和種は、水田の耕起、農作物の運搬など役 用を主たる目的として飼養されていたが、1960年頃から農作業の機械化およ び牛肉消費の増大に伴い、役用から肉専用種への転換が図られ、短期間のうち にその転換に成功した。比較的小規模な品種でありながら、他品種から遺伝子 を導入することなしに、短期間のうちにこのような一大転換に成功した品種は、 世界的に見ても少ないはずである。この成功には、品種が分化構造を呈してお り、肉質や増体に優れた遺伝子を持ついくつかの分集団(系統)が品種内に存 在したことが大きく寄与したものと考えられる。過去の改良過程において遺伝 的多様性が果たした役割から考えると、他品種からの遺伝子の導入がない閉じ られた繁殖集団を形成しつつ、将来の改良目標の変更に対応するための遺伝的 な可塑性を維持するには、長期的な視野に立った改良方針の樹立が急務である と考えられる。 諸外国の品種がいずれも100前後の有効な大きさを保ってきたこと、そして 黒毛和種は他品種からの移入のない閉鎖集団であることを考え合わせると、少 なくとも100以上の有効な大きさは確保すべきであろう。今後、集団の有効な 大きさを定期的にモニタリングするシステムを確立しておくことも重要である と考える。 現在、和牛の遺伝的評価は枝肉形質を中心に行われているが、他の重要な能 力である繁殖性や母性能力についての評価も実施の準備が整いつつある。この ような総合的な能力評価が可能になったときに、現在の肉質に極端に傾斜した 種雄牛の選定が少しでも緩和され、集団の有効な大きさが拡大されることに期 待したい。和牛において「遺伝的多様性を確保する」ために最も有効な方策は、 「改良目標に多様性を持たせる」ことであると考える。 注:詳しい推定方法の解説は、「和牛誌」2000年1月号(全国和牛登録協会) を参照していただきたい。