栄養/健康
牛肉・牛肉エキス摂取による骨格筋特性の栄養応答
―牛肉摂取による身体能力の特性と向上を目指して―
九州大学大学院農学研究院 教授 池内 義秀
栄養/健康牛肉・牛肉エキス摂取による骨格筋特性の栄養応答
|
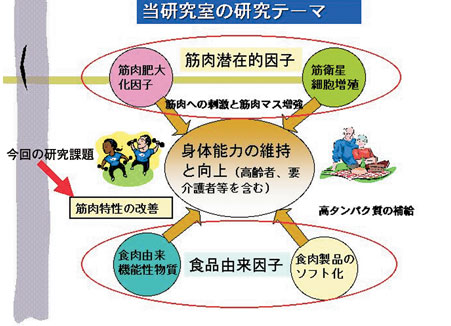
はじめに 高齢者の増加は、医療費、介護費などの社会的コストの増大だけにとどまらず、高齢者個人と高齢者の生活を世話する家族などの負担の増大となり、それ自体が深刻な社会問題を引き起こしている。介護保険などの社会福祉制度の充実は当然ではあるが、一方で高齢者が虚弱化して生活活動能力を喪失し、寝たきり、要介護状態に陥るのを出来るだけ防止するためにも、高齢者向けの運動と併せて食事法の改善が有効であると提唱されている。われわれも、図1に示しているように、筋肉に潜在的に存在する能力、および食品とりわけ食肉由来の因子を利用して高齢者や要介護者の身体能力の維持、向上を目指す取り組みを行っている。
|
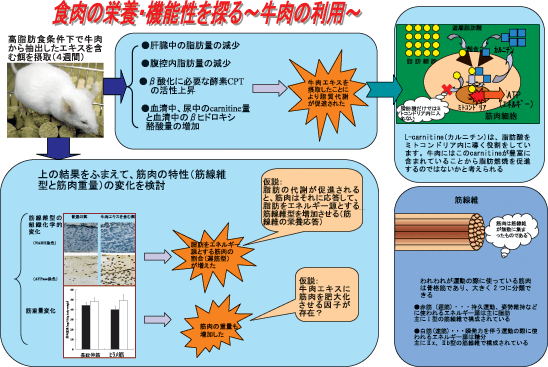 図2 牛肉摂取がラットの筋肉特性に及ぼす影響 |
筋肉には収縮速度が遅く(遅筋)、有酸素系のエネルギー代謝を主に行うI型筋線維(slow-twitch,oxidative)と収縮速度が速く(速筋)、解糖系の代謝を主に行うIIa、IIx、IIb型筋線維(fast-twitch,glycolytic)などに分けられる(動物種によって分類の仕方は異なる)。筋肉の色素成分であるミオグロビンの量の差による色の違いから、遅筋は赤筋、速筋は白筋とも呼ばれている。筋線維型は持久的トレーニングにより速筋型から遅筋型へ、筋力トレーニングにより速筋型から中間型へ、無重力などの除負荷や低温刺激により遅筋型から速筋型へと変化することが知られているが、この筋線維型の変化はカルシニュリンという脱リン酸化酵素により調節されていると考えられている。その機構は運動や神経刺激により細胞内カルシウム濃度が上昇すると、まずカルシニュリンが活性化され、タンパク質NFATが脱リン酸化されて、それが核内でMEF-2という転写因子との相互作用により遅筋線維型筋線維タンパク質の発現が上昇するというものである。
牛肉エキス摂取が長趾伸筋線維組成(速筋)を変化させた理由については目下不明である。しかしながら、代謝関連遺伝子のmRNAが摂取食餌の質と量といった栄養条件によって制御を受けていることが知られていること、また、骨格筋は全身の糖代謝、脂肪代謝のうちの大きな割合を占め、各筋線維型によりその代謝能力が異なることなどから、牛肉エキスに含まれる成分が何らかの生理的機能を有し、これが長趾伸筋細胞に作用して最終的に有酸素系代謝(酸化的リン酸化)活性の高い遅筋型筋線維を量的に増加させた可能性がある。ここで候補となり得るのは、牛肉エキスに高濃度に含まれ、ミトコンドリア内膜に長鎖脂肪酸を輸送する担体として働き、脂肪酸のβ酸化に重要な役割を演じているカルニチンである(図2)。L-カルニチン(4-trimethylamino-3-hydroxy butyric acid)は骨格筋など生体内にもともと存在するジペプチドで、食事と体内での生合成により賄われている。体内ではリジンとメチオニンにより主に肝臓と腎臓で生合成される。一般の食品では動物性食品に多く含まれており、特に牛肉や羊肉に多い。L-カルニチンが最初に発見されたのは1905年で、肉エキスから分離された。そして1955年には脂肪酸の酸化に重要な働きをもつこと、1959年には肝臓での遊離脂肪酸の酸化に必要であることが明らかにされた。1973年にはヒトでカルニチン欠乏症が発見され、以来カルニチンの生理活性作用に関する研究が活発に進められている。実際カルニチン摂取により脂質代謝が亢進されるという報告が数多くある。
以上、骨格筋の筋線維型は上述の通りエネルギー代謝特性に依存していることから、骨格筋細胞は運動負荷という物理的刺激だけではなく、栄養要因によっても最適に応答する能力を有しているのではないかと予想している。
骨格筋の肥大には、構成している筋線維(筋細胞)の数の増加と大きさの増大が関与する。筋重量の実質的な増大は筋原線維を構成するタンパク質の合成が増大し、分解が抑制されているか、あるいは分解は亢進しないがタンパク質の蓄積が起こるためと考えられる。一方、骨格筋には、分化の過程でコミットメントしたが増殖しないまま基底膜に休止状態で入っている増殖可能な幹細胞(筋衛星細胞)が多く存在し、それが何らかの刺激を受けて増殖し、新しい筋細胞をつくる。それ故、筋衛星細胞の活性化も筋肥大には重要である。生体では筋細胞自身が分泌するインシュリン様成長因子(IGF-1)、神経由来のS-マイオトロフィン(当研究室で精製された筋肥大化因子)、成長ホルモン、ステロイドホルモン、あるいは外来性の栄養素などによって筋細胞や筋衛星細胞が刺激され、成長・肥大化する。実際、牛肉エキスにこのような筋肉肥大化に関与する成分が含まれているのか、または食肉由来の消化産物を含めて未知の因子が存在するのかどうか現時点では不明である。
今回の研究では、より脂質代謝の亢進を期待して、カルニチンと同じく動物性食品中(特に反すう動物)に含まれ、体脂肪低減効果があるといわれている共役リノール酸(以下CLA)をカルニチンと同時摂取させた。CLAは脂肪酸の一種であるリノール酸の位置および構造異性体の総称で反すう動物由来の食品中に含まれている。今回供試したCLAは、2つの主要な異性体をほぼ同量含んだものであった。その生理機能としては、カルニチンが脂肪酸をミトコンドリア内に運ぶための担体として働くのに対して、CLAは体脂肪の脂肪とグリセリンへの分解を促進すると言われている。
7週齢Wistar系雄ラットをセアック吉冨(株)より購入し、1週間市販飼料(CRF-1 : 日本チャールズリバー(株))で予備飼育後、0.5%カルニチン摂取区と非摂食区に分け、さらに共役リノール酸とカルニチンの相乗効果を調べるために、それぞれ1%CLA摂取区・非摂取区の計4グループに分け、高脂肪食条件下で4週間飼育した。飼料は1匹当たり1日30グラム供与し、これを自由摂取させて食べ残しを計量し、摂取量を求めた。水は自由摂取とした。飼育は各群ステンレス製の個別ケージ(20×25×17センチメートル)で室温20℃、60%湿度、12時間照明の空調動物室内(九州大学生物環境調節センター:バイオトロン)にて行った。飼育期間最終日に一晩絶食させ、エーテル麻酔下でと殺し、血液を採取後肝臓・腹腔内脂肪・脚部筋肉を摘出して重量を測定した。摘出した筋肉は重量を測定したのち直ちに冷アセトンによりいったん凍結し、SDSサンプル作製まで−80℃のディープフリーザーで保存した。電気泳動後CBB染色を行い、画像処理ソフトNIHimageを使ってミオシンの4つのアイソフォームの割合を求めた。泳動条件は、8%ポリアクリルアミドゲルで8℃,100V, 20hとした。
筋芽細胞株L6を6φのDishに播種し、筋管細胞へと分化させた。その後、牛肉エキスあるいはカルニチンを含む培地(DMEM+10%馬血清)を添加して、72時間培養した。培養終了後、顕微鏡を用いて、各Dishの各視野における筋管細胞の最大直径を計測した。筋線維型の変化を調べるために、培養筋細胞をlysate bufferで溶解後、遅筋型ミオシンモノクローナル抗体を用いて、ウエスタンブロッティングを行った(チューブリン抗体を内部標準に用いた)。また、筋肥大を引き起こす代表的な因子であるIGF-Iが牛肉エキスに存在するのか、またそれが筋肥大に関与しているかを調べるために、その実験区とは別に、上記の実験条件にIGF-I中和抗体添加区を設けた。
カルニチンおよびCLA摂取において飼料摂取量に差がないにもかかわらず、腹腔内脂肪量・腹腔脂肪率の低下が認められた(図3)。
さらに、血中カルニチン量は、カルニチン摂取の2グループにおいて有意に増加した。カルニチンは経口投与では50〜80%が吸収されると言われているが、本実験でもエキス由来のカルニチンが体内に十分吸収されたことがわかった。また、肝臓と同様血清中GLU・TG・TCHOはカルニチン・CLA摂取により減少した。以上の結果から、期待したほどCLA摂取との相乗効果は見られなかったものの、カルニチン摂取はラット体内の脂質代謝を亢進することが再確認された(図 3)。
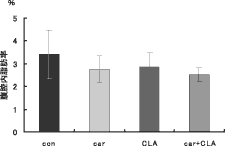
図3 カルニチンおよびCLA摂取がラット腹腔脂肪率に及ぼす影響
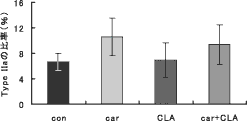 図4 カルニチンおよびCLA摂取がラットEDL筋の筋線維型に及ぼす影響 |
ラットの長趾伸筋(EDL:Extensor digitorum longus)は90%以上が速筋(II型)、ヒラメ筋(Soleus)は90%以上が遅筋(I型)であることが知られている。図4で示したように、EDLでは牛肉エキス摂取動物実験結果と同様、カルニチン単独摂取でも酸化的代謝を行う筋線維の割合が増加する傾向が認められた。筋肉培養細胞を用いた実験でも、カルニチン投与により遅筋型の筋線維が増加することが確かめられた(図5)。
一方、もともと有酸素系の代謝を行うヒラメ筋では筋線維型の変化は見られなかった。またCLA摂取区でもそのような筋腺維型の変化は見られなかった。

図5 カルニチン投与が骨格筋由来L6培養細胞の筋線維型に及ぼす影響
72時間培養した後骨格筋slow type myosin monoclonal antibodyを用い
てウエスタンブロッティングを行った。
牛肉エキス摂取はラットの筋肉量を増加させたが(図6)、カルニチン単独ではそのような効果はなかった。そこで、牛肉エキスが筋肉培養細胞の筋管を大きくするかどうか調べたところ、図7に示したように、牛肉エキスは濃度に依存して培養筋細胞の筋管肥大を引き起こすことがわかった。次に、筋肥大に関与する因子、IGF-1が牛肉エキスに含まれている可能性があるので、IGF-1の中和抗体を牛肉エキスとともに培養細胞に投与した。その結果、十分量の抗体の投与にもかかわらず、牛肉エキスにより筋管細胞の最大直径が有意に増加した(データ未発表)。以上の結果から、牛肉エキス中には筋管細胞の肥大を誘導する因子が含まれていること、また、それはIGF-Iではなく、またIGF-Iの発現を誘導する因子でもないことが明らかになった。
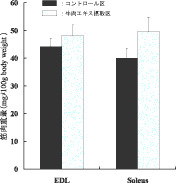 図6 牛肉エキス摂取によるラット長趾伸筋(EDL)およびヒラメ筋の重量変化 |
 図7 牛肉エキス投与がL6培養細胞の筋管の肥大に及ぼす影響 培養開始直後(0hr)と培養72時間後のL6筋細胞の顕微鏡写真 |
以上の結果より、カルニチンとCLA摂取による相乗効果は見られなかったものの、脂質代謝の亢進がみられ、さらにカルニチン摂取という栄養効果により筋線維型の変化が起こり得ることが示唆された。おそらく、カルニチン摂取によりβ-酸化が亢進されたことで何らかの筋線維型変換誘導因子が刺激され、それが遅筋型の筋線維型の発現を促したのではないかと思われる。つまり、骨格筋細胞は運動・神経刺激という物理的刺激だけでなく、栄養状態によっても最適に応答する能力を有していると考えられる。一方、牛肉エキスには骨格筋の肥大化を引き起こす成分が含まれていることが、生体レベルおよび細胞培養実験で裏付けられた。
筋線維型の変化や筋肉肥大は多くの要因が複雑に関係して起こると考えられるため、栄養条件による影響(栄養応答)も、その要因の一つに過ぎないかもしれない。今後、栄養摂取と筋肉の肥大、筋線維型変化との関係、メカニズムを明らかにするためには、分子生物学的手法による脂質代謝関連酵素遺伝子群や筋線維型にかかわる遺伝子群の網羅的解析あるいはプロテオーム解析、また筋肥大化成分の特定を行う必要があると思われる。
以上、運動を伴わず牛肉エキスという栄養摂取のみで骨格筋の筋量や筋線維型が変化、特に持久型の筋線維が増加するという事実は、高齢者の身体能力維持あるいは予後の体力回復、また、アスリートにとっても疲労回復と身体能力の維持・向上に役立つものと期待される。
元のページに戻る